
Das Meselson-Stahl-Experiment gehört zu den saubersten Belegen in der Molekularbiologie: Es zeigt, wie DNA sich verdoppelt und warum jede neue Doppelhelix aus einem alten und einem neuen Strang besteht. Für das Verständnis von Vererbung ist das kein Detail, sondern der Kern der Sache. Ich gehe hier deshalb Schritt für Schritt durch die Fragestellung, die Methode mit schwerem und leichtem Stickstoff, die beobachteten Banden und die Konsequenzen für die Biologie.
Die wichtigsten Punkte auf einen Blick
- Der Versuch prüfte, ob DNA konservativ, semikonservativ oder dispersiv repliziert wird.
- Verwendet wurden ^15N- und ^14N-Stickstoff sowie CsCl-Dichtegradientenzentrifugation.
- Nach einer Replikationsrunde erschien nur eine Zwischenbande, nach der zweiten eine Zwischenbande und eine leichte Bande.
- Damit fielen das konservative und das dispersive Modell weg; übrig blieb die semikonservative Replikation.
- Die Methode ist bis heute ein Lehrbuchbeispiel dafür, wie man eine biologische Hypothese experimentell sauber prüft.
Welche Frage der Versuch überhaupt klären sollte
Ich halte diesen Versuch vor allem deshalb für stark, weil er keine diffuse Theorie testen wollte, sondern eine sehr konkrete Frage: Was passiert mit der alten DNA beim Kopieren? In den 1950er-Jahren standen drei Modelle nebeneinander, und jedes machte andere Vorhersagen für die Nachkommenschaft einer DNA-Doppelhelix.
Die drei Möglichkeiten lassen sich knapp so unterscheiden: Entweder bleibt der alte Doppelstrang vollständig erhalten und eine komplett neue DNA kommt hinzu, oder jeder Strang dient als Vorlage für einen neuen Partner, oder die alte und die neue DNA werden in kleinen Stücken gemischt. Genau diese Unterschiede sind entscheidend, weil sie direkt sagen, wie man die Erbinformation versteht.
| Modell | Vorhersage nach 1 Replikationsrunde | Vorhersage nach 2 Replikationsrunden | Was das bedeuten würde |
|---|---|---|---|
| Konservativ | Eine schwere und eine leichte Bande | Schwere Bande bleibt, leichte Bande nimmt zu | Der elterliche Doppelstrang bleibt komplett erhalten |
| Semikonservativ | Nur eine Zwischenbande | Zwischenbande und leichte Bande | Jedes neue Molekül enthält einen alten und einen neuen Strang |
| Dispersiv | Nur eine Zwischenbande | Weiterhin nur eine Bande, die sich langsam verschiebt | Alte und neue DNA wären über das ganze Molekül verteilt gemischt |
Genau an dieser Stelle wird das Experiment interessant: Nicht die Theorie an sich war das Problem, sondern die Frage, welche Vorhersage sich im Labor wirklich messen lässt. Dafür brauchte es eine Methode, die kleinste Dichteunterschiede sichtbar macht.
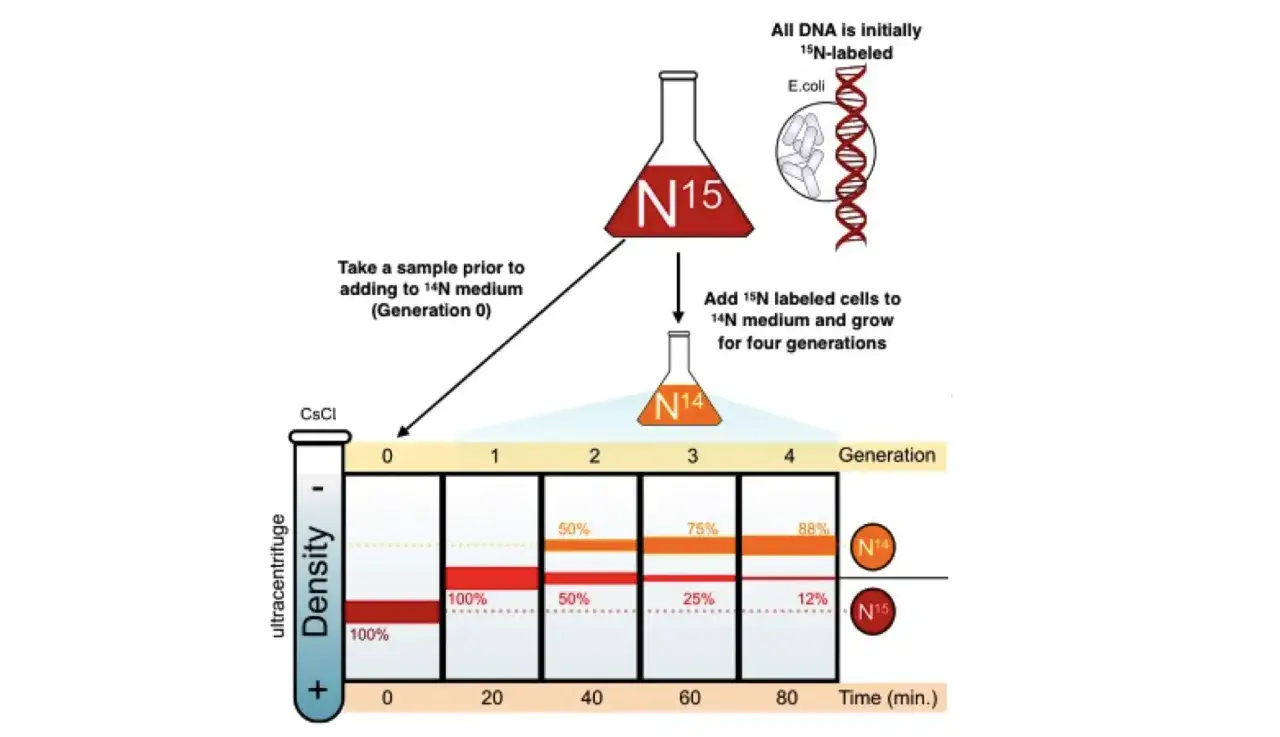
Wie Meselson und Stahl die DNA sichtbar machten
Die Versuchsidee war elegant und technisch anspruchsvoll zugleich. Meselson und Stahl züchteten E. coli zunächst in einem Medium mit schwerem Stickstoff (^15N), damit sich das Isotop in die DNA einbaute. Danach wechselten sie die Bakterien in ein Medium mit leichtem Stickstoff (^14N), sodass ab diesem Moment neue DNA mit dem leichteren Isotop aufgebaut wurde.
Ich würde diesen Teil immer in drei Schritten erklären, weil er sonst unnötig kompliziert wirkt:
- Die Bakterien wurden über viele Generationen im schweren Medium gehalten, damit ihre DNA vollständig markiert war.
- Dann kam der Wechsel auf leichtes Stickstoffmedium, und die Zellen replizierten ihre DNA weiter.
- Die isolierte DNA wurde in einer CsCl-Lösung zentrifugiert, sodass sich Moleküle nach ihrer Auftriebsdichte in Banden anordnen konnten.
Der wichtige Punkt ist dabei nicht das Gewicht im alltagssprachlichen Sinn, sondern die Dichte im Gradient. Die DNA wandert an die Stelle, an der ihre eigene Dichte und die Dichte der Lösung im Gleichgewicht sind. So lassen sich Moleküle mit unterschiedlicher Isotopenzusammensetzung voneinander trennen.
Gerade diese Kombination aus Isotopenmarkierung und Dichtegradientenzentrifugation machte den Versuch so überzeugend. Als Nächstes zählt nur noch, welche Banden tatsächlich erschienen.
Was die Banden im Röhrchen bewiesen
Nach der ersten Replikationsrunde sahen Meselson und Stahl keine zwei getrennten Banden, sondern eine einzige Bande genau zwischen schwer und leicht. Das ist der entscheidende Befund, denn ein konservatives Modell hätte sofort zwei Banden erzeugt: eine schwere für die unveränderte Eltern-DNA und eine leichte für die neu gebildete DNA.
Nach einer zweiten Replikationsrunde trat dann das Muster auf, das man heute aus jedem guten Lehrbuch kennt: eine Zwischenbande und eine leichte Bande. Das passt sauber zur semikonservativen Replikation, weil sich die Hybridmoleküle weiter verdünnen, während neue, vollständig leichte DNA entsteht.
- Eine Zwischenbande nach einer Runde schließt das konservative Modell aus.
- Eine Zwischenbande plus eine leichte Bande nach zwei Runden passt zur semikonservativen Replikation.
- Ein dauerhaft einzelnes Zwischenband hätte für ein dispersives Modell gesprochen, wurde aber nicht beobachtet.
Ich würde die Schlussfolgerung so zuspitzen: Die alte DNA bleibt nicht als geschlossener Block erhalten, sondern jeder Strang dient als Vorlage für einen neuen Partner. Damit war die Watson-Crick-Idee nicht nur plausibel, sondern experimentell sehr stark abgesichert.
Gerade deshalb wurde das Ergebnis so wichtig für die gesamte Molekularbiologie.
Warum dieses Ergebnis die Molekularbiologie verändert hat
Der eigentliche Wert des Experiments liegt nicht nur darin, dass eine alte Streitfrage gelöst wurde. Es hat gezeigt, dass DNA-Vererbung auf einem präzisen Vorlagenprinzip beruht: Jeder Elternstrang trägt Information, an die komplementär ein neuer Strang gebaut wird. Das ist die logische Brücke zwischen Struktur und Funktion.
Für die Biologie war das ein Durchbruch, weil damit klar wurde, warum genetische Information so zuverlässig kopiert werden kann und gleichzeitig trotzdem Veränderung möglich bleibt. Fehler in der Replikation können dann als Mutationen auftreten, und genau diese Mischung aus Stabilität und Variabilität ist biologisch entscheidend.
Auch im Unterricht oder Studium ist das Thema bis heute zentral, weil es gleich mehrere Grundideen zusammenführt: Basenpaarung, Komplementarität, Vorlageprinzip und experimentelle Kontrolle. Ich finde das besonders lehrreich, weil man hier sieht, wie eine gute biologische Frage mit einer sauberen Methode beantwortet wird, statt mit bloßer Plausibilität.
Trotzdem wird der Versuch oft zu grob vereinfacht, und genau da entstehen die typischen Missverständnisse.
Wo Lernende und Lehrbücher oft zu grob werden
Der Versuch beweist die Art der Replikation, nicht gleich die komplette Enzymmaschinerie dahinter. Er sagt also nicht, welche Polymerase in welchem Schritt arbeitet, wie der Leit- und Folgestrang organisiert ist oder wie der Replikationsgabel-Komplex genau aufgebaut ist. Diese Details kamen erst später durch andere Experimente hinzu.
Ein zweites Missverständnis betrifft die Zwischenbande. Sie bedeutet nicht, dass sich die DNA in der Mitte „halbiert“ hätte, sondern dass Moleküle mit gemischter Isotopenzusammensetzung eine Dichte zwischen schwer und leicht besitzen. Das ist ein physikalischer Messwert, kein symbolisches Bild.
Außerdem lohnt sich eine kleine methodische Vorsicht: Das Experiment funktioniert nur so gut, weil die Banden im CsCl-Gradienten wirklich trennscharf sind und die Ausgangskultur sauber markiert war. Schlechte Trennung, Kontaminationen oder unklare Generationenzahlen würden die Interpretation schnell verwischen.
- Das Experiment beantwortet die Frage nach dem Replikationsmodus, nicht die nach allen beteiligten Enzymen.
- Die Zwischenbande ist ein Dichteeffekt, kein Zeichen für eine „halb fertige“ DNA im umgangssprachlichen Sinn.
- Die Aussage ist robust, aber sie hängt an sauberer Isotopenmarkierung und präziser Zentrifugation.
- Das semikonservative Prinzip gilt breit, auch wenn sich die Organisation der DNA in verschiedenen Organismen unterscheidet.
Genau diese Klarheit ist der Grund, warum der Versuch bis heute so oft erklärt wird und trotzdem nicht banal wirkt.
Was man sich aus dem Versuch bis heute merken sollte
Wenn ich den Inhalt auf drei Sätze kürze, dann so: Erstens war die Frage nach dem Replikationsmodus offen. Zweitens lieferten ^15N und ^14N zusammen mit der CsCl-Dichtegradientenzentrifugation einen messbaren Unterschied zwischen den Modellen. Drittens zeigte die Kombination aus Zwischenbande und späterer Aufspaltung in Zwischen- und leichte Bande eindeutig die semikonservative Replikation.
- Vorlageprinzip heißt: Jeder alte Strang dient als Muster für einen neuen.
- Semikonservativ heißt: Jedes Tochtermolekül behält einen Elternstrang.
- Experimentell elegant heißt hier: Eine einfache Vorhersage, ein klarer Messwert, eine eindeutige Schlussfolgerung.
Wer den Versuch verstanden hat, hat nicht nur ein Stück Wissenschaftsgeschichte im Kopf, sondern auch ein sehr gutes Gefühl dafür, wie Biologie echte Hypothesen prüft. Genau diese Denkweise ist der bleibende Wert des Meselson-Stahl-Experiments.