
Die Reifung von RNA entscheidet darüber, ob aus einem frisch transkribierten Vorläufer eine nutzbare Botschaft, ein stabiles Adaptermolekül oder ein funktionsfähiger Ribosomenbaustein wird. Ich ordne das Thema deshalb in drei Ebenen: mRNA, tRNA und rRNA. Dazu zeige ich, welche Schritte in der Zelle nacheinander greifen, warum Spleißen und chemische Modifikationen so viel bewirken und wo Fehler in der Praxis biologisch sichtbar werden.
Die wichtigsten Punkte zur RNA-Reifung auf einen Blick
- Die meisten eukaryotischen RNAs werden erst nach der Transkription funktionsfähig.
- Bei mRNA sind 5'-Cap, Spleißen und Polyadenylierung die drei Kernschritte.
- Alternatives Spleißen erzeugt aus einem Gen mehrere RNA- und Proteinvarianten.
- tRNA und rRNA werden anders verarbeitet, enthalten aber ebenfalls viele Modifikationen.
- Fehler in der Prozessierung können zu instabilen Transkripten, falschen Proteinen und Krankheiten führen.
Was RNA-Prozessierung in der Zelle eigentlich bedeutet
Wenn ich RNA-Prozessierung knapp definieren müsste, würde ich sagen: Es ist die Gesamtheit aller Schritte, die aus einer frisch gebildeten Vorstufe eine reife, funktionierende RNA machen. Bei Eukaryoten passiert das meist im Zellkern und oft co-transkriptionell, also noch während die RNA-Polymerase den Strang überhaupt erst synthetisiert. Diese Nähe von Transkription und Reifung ist kein Nebeneffekt, sondern ein Kontrollprinzip.
Die wichtige Unterscheidung lautet deshalb nicht nur „RNA oder DNA“, sondern „Vorstufe oder Endprodukt“. Eine prä-mRNA trägt noch Introns, unvollständige Enden und oft noch keine Signale für Export oder Translation. Erst die Prozessierung macht daraus eine RNA, die der Zellkern als reif akzeptiert und die im Cytoplasma sinnvoll gelesen werden kann. Genau hier liegt die praktische Bedeutung: Nicht jede transkribierte RNA ist automatisch biologisch aktiv.
Für die Biologie ist das deshalb so relevant, weil Prozessierung mehrere Aufgaben gleichzeitig erfüllt: Sie schützt vor Abbau, legt Leserahmen fest, erhöht die Übersetzbarkeit und schafft regulierbare Varianten. Der nächste Schritt ist daher fast immer die Frage, wie genau die einzelne Vorstufe bearbeitet wird - und bei der mRNA ist das besonders gut untersucht.
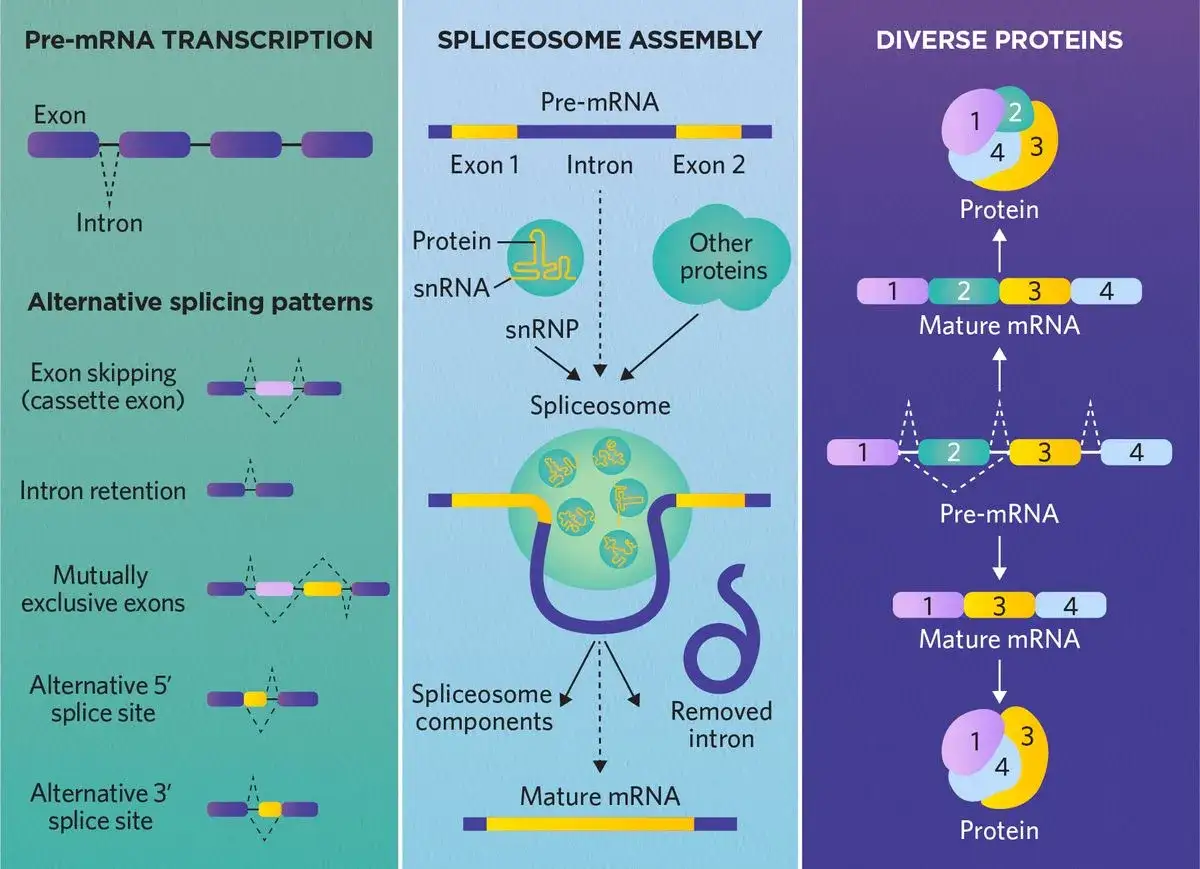
Wie aus prä-mRNA eine reife mRNA wird
Die klassische mRNA-Reifung besteht aus drei eng gekoppelten Schritten: 5'-Capping, Spleißen und 3'-Endverarbeitung. Ich finde diese Abfolge didaktisch so wichtig, weil sie zeigt, dass die Zelle nicht einfach „abschneidet“, sondern gezielt markiert, sortiert und stabilisiert. Jede Änderung erfüllt eine eigene Funktion.
Das 5'-Cap schützt und markiert das Transkript
Am 5'-Ende wird sehr früh ein 7-Methylguanosin-Cap angefügt. Diese Struktur schützt die RNA vor Abbau, hilft beim Export aus dem Kern und erleichtert später die Erkennung durch das Translationssystem. In der Praxis ist das Cap also nicht bloß eine chemische Deko, sondern ein Funktionssignal. Ohne es verliert die Zelle ein Stück Kontrolle über die Lebensdauer der mRNA.
Das Spleißen entfernt Introns präzise
Beim Spleißen werden Introns entfernt und Exons miteinander verknüpft. Das erledigt das Spliceosom, ein großer Komplex aus Proteinen und kleinen nukleären RNAs, die man als snRNAs bezeichnet. Besonders wichtig sind dabei die Erkennungsstellen am 5'-Spleißort, am Branch-Point und am 3'-Spleißort. Wird eine dieser Stellen falsch erkannt, kann das Leseraster kippen oder ein Protein völlig anders ausfallen als vorgesehen.
Mechanistisch läuft das in zwei Reaktionsschritten ab: Zuerst entsteht eine Schleifenstruktur, das sogenannte Lariat, dann werden die Exons ligiert. Ich halte diese Lariat-Form für einen der anschaulichsten Punkte der molekularen Biologie, weil sie gut zeigt, dass RNA nicht nur Information trägt, sondern selbst strukturell und katalytisch aktiv organisiert wird.
Die 3'-Endverarbeitung stabilisiert die Botschaft
Am anderen Ende der RNA wird geschnitten und meist ein Poly-A-Schwanz angefügt. In vielen mRNAs umfasst er ungefähr 100 bis 250 Adeninreste; oft wird grob mit rund 200 gearbeitet. Diese Verlängerung stabilisiert das Transkript, beeinflusst den Export und wirkt auf die spätere Translation. Ein bekanntes Signal in vielen Säugerzellen ist die Sequenz AAUAAA, die der Zelle hilft, die passende Schnittstelle zu finden.
Wichtig ist für mich hier der Gedanke der Kopplung: Capping, Spleißen und Polyadenylierung sind keine isolierten Inseln. Sie beeinflussen sich gegenseitig und hängen mit der Polymerase II zusammen, die den gesamten Prozess koordiniert. Genau diese Verzahnung erklärt, warum Fehler an einer Stelle oft Folgen an mehreren anderen haben.
Warum alternatives Spleißen aus einem Gen mehrere Varianten macht
Alternatives Spleißen ist einer der elegantesten Mechanismen der Genregulation. Aus derselben prä-mRNA können unterschiedliche Exonkombinationen entstehen, und damit verschiedene mRNAs - oft mit deutlich unterschiedlicher Funktion. Für die Zelle ist das ein massiver Effizienzgewinn: Mehr biologische Vielfalt aus weniger Genen.
Die häufigsten Muster lassen sich gut unterscheiden:
- Exon-Skipping, bei dem ein Exon übersprungen wird.
- Alternatives 5'- oder 3'-Spleißen, bei dem unterschiedliche Spleißstellen gewählt werden.
- Wechselseitig ausschließende Exons, bei denen nur eines von zwei Exons eingebaut wird.
- Intron-Retention, bei der ein Intron im Transkript verbleibt.
Reguliert wird das durch RNA-bindende Proteine und sequenzielle Signale in der prä-mRNA. SR-Proteine fördern oft bestimmte Spleißentscheidungen, während hnRNPs andere Muster begünstigen oder bremsen. Das ist nicht nur akademisch interessant: Gewebe, Entwicklungsstadien und Stressbedingungen nutzen genau diese Mechanik, um Proteine an ihre jeweilige Aufgabe anzupassen.
Ich sehe hier auch die praktische Grenze vieler vereinfachter Darstellungen. Alternatives Spleißen erzeugt nicht automatisch „bessere“ Varianten. Manchmal entsteht nur eine andere, und nicht immer funktionelle, Isoform. Der biologische Effekt hängt davon ab, welche Domänen im Protein erhalten bleiben, wo das Protein wirken soll und wie streng die Zelle das Ergebnis kontrolliert. Von dort ist es nur ein Schritt zu den anderen RNA-Klassen, die ebenfalls intensiv verarbeitet werden.
tRNA und rRNA werden anders, aber ebenso gründlich verarbeitet
Neben der mRNA sind vor allem tRNA und rRNA klassische Beispiele für RNA-Reifung. Beide werden aus Vorläufern erzeugt, aber ihr Weg ist nicht identisch mit der mRNA. Gerade daran sieht man, wie flexibel Zellen mit derselben Grundchemie umgehen.
| RNA-Typ | Typische Vorstufe | Wichtige Verarbeitung | Biologischer Zweck |
|---|---|---|---|
| mRNA | prä-mRNA | 5'-Cap, Spleißen, Polyadenylierung | Transportierbare Vorlage für die Proteinsynthese |
| tRNA | prä-tRNA | Trimmen der Enden, Anfügen von CCA, viele Basenmodifikationen | Adapter zwischen Codon und Aminosäure |
| rRNA | prä-rRNA | Spaltung, Faltung, chemische Modifikationen, Einbau in Ribosomen | Strukturelle und katalytische Grundlage des Ribosoms |
tRNA braucht präzises Trimmen und viele Modifikationen
tRNAs werden an beiden Enden bearbeitet, und am 3'-Ende kommt häufig die Sequenz CCA hinzu, falls sie nicht bereits im Gen kodiert ist. Diese Endsequenz ist für die Anlagerung der Aminosäure unverzichtbar. An der 5'-Seite spielt die RNase P eine zentrale Rolle, ein Enzymkomplex, der historisch auch deshalb spannend ist, weil seine RNA-Komponente katalytisch aktiv sein kann.
Hinzu kommen zahlreiche chemische Veränderungen einzelner Basen. Grob gesagt ist bei einer reifen tRNA ungefähr jede zehnte Base chemisch verändert. Das klingt nach einer Kleinigkeit, hat aber große Folgen für Stabilität, Erkennung und Genauigkeit bei der Translation. Wer verstehen will, warum tRNAs so zuverlässig arbeiten, kommt an diesen Modifikationen nicht vorbei.
Lesen Sie auch: Fotosynthese-Gleichung verstehen - Mehr als nur CO2 + H2O
rRNA wird geschnitten, gefaltet und gezielt verändert
Auch rRNA entsteht nicht einfach in fertiger Form. Große Vorläufer werden geschnitten, chemisch modifiziert und gemeinsam mit Ribosomenproteinen zu funktionsfähigen Partikeln aufgebaut. In Eukaryoten leiten snoRNAs viele dieser Modifikationen, indem sie per Basenpaarung die Zielstellen im Vorläufer erkennen. Besonders häufig sind dabei Methylierungen und Pseudouridin-Bildungen.
Ein wichtiger Sonderfall ist die 5S-rRNA, die separat transkribiert wird und nicht denselben Reifungsweg wie die großen rRNA-Vorstufen durchläuft. Genau solche Unterschiede sind für Lernende oft verwirrend, aber biologisch aufschlussreich: Nicht jede RNA folgt demselben Reifungsmodell, obwohl das Ziel immer dasselbe bleibt - Funktionstüchtigkeit. Von hier aus ist der Schritt zu chemischen Feinjustierungen und RNA-Editing naheliegend.
Chemische Modifikationen und RNA-Editing verändern die Funktion
Ich würde RNA-Reifung unvollständig nennen, wenn man nur an Schneiden und Zusammenfügen denkt. Chemische Modifikationen sind ein zweiter großer Layer, der die Funktionsweise der RNA fein abstimmt. Dazu gehören unter anderem 2'-O-Methylierungen, Pseudouridylierungen, m6A-Markierungen und in manchen Kontexten auch RNA-Editing, etwa A-zu-I-Umwandlungen.
Der entscheidende Punkt ist: Diese Veränderungen verlängern oder verkürzen eine RNA nicht zwangsläufig, sondern verändern ihre Eigenschaften. Sie können die Faltung stabilisieren, die Lebensdauer beeinflussen, die Bindung von Proteinen erleichtern oder erschweren und die Translation mitsteuern. Gerade bei mRNA ist das biologisch relevant, weil zwei Transkripte mit identischer Sequenz funktionell nicht gleich behandelt werden müssen, wenn ihr Modifikationsmuster anders ist.
Bei rRNA und tRNA sind solche Modifikationen besonders dicht. Dort geht es oft um Präzision im Millisekunden- bis Minutenbereich: korrekte Geometrie, stabile Interaktion und fehlerarme Proteinsynthese. In der Forschung ist das Feld inzwischen so groß, dass man Modifikationen nicht mehr als Randnotiz behandelt, sondern als eigenen Regulationscode. Das macht die Sache komplexer, aber auch biologisch viel realistischer.
Damit ist die eigentliche Frage schnell erreicht: Was passiert, wenn diese Reifung nicht sauber läuft?
Wo Fehler in der Prozessierung sichtbar werden
Fehler in der RNA-Reifung bleiben selten folgenlos. Eine falsch gespleißte mRNA kann ein verschobenes Leseraster erzeugen, ein vorzeitiges Stoppsignal enthalten oder ein wichtiges Proteindomänensegment verlieren. Das Resultat ist dann nicht einfach „ein bisschen weniger effizient“, sondern häufig ein ganz anderes oder gar kein funktionsfähiges Protein.
Typische Fehlerquellen sind:
- Mutationen an Spleißstellen oder Branch-Points.
- Defekte in Spliceosom-Komponenten.
- Störungen beim Capping oder bei der Polyadenylierung.
- Unvollständige Reifung von tRNA- oder rRNA-Vorläufern.
- Überlastete Qualitätskontrollen bei Stress oder Krankheit.
Die Zelle reagiert darauf mit Überwachungssystemen. Unreife oder fehlerhafte Transkripte können im Zellkern zurückgehalten und abgebaut werden. Auch die sogenannte nonsense-mediated decay ist wichtig: Sie erkennt mRNAs mit vorzeitigem Stopcodon und verhindert so oft die Bildung schädlicher Proteine. Für mich ist das ein gutes Beispiel dafür, dass RNA-Reifung und Qualitätskontrolle keine getrennten Themen sind, sondern zusammengehören.
Genau hier wird die Relevanz für Medizin und Forschung greifbar. Viele genetische Erkrankungen, aber auch bestimmte Tumorveränderungen, hängen nicht nur an der DNA-Sequenz selbst, sondern daran, wie die Zelle diese Information verarbeitet. Das führt direkt zu der Frage, was man sich vom Thema praktisch merken sollte.
Woran sich gute RNA-Reifung erkennen lässt
Wenn ich das Thema auf drei Sätze herunterbrechen müsste, dann wären es diese: Erstens wird RNA in Eukaryoten meist vor ihrer Nutzung bearbeitet. Zweitens entscheidet die Art der Bearbeitung oft über Stabilität, Vielfalt und Funktion. Drittens sind Fehler in diesem Prozess kein Randproblem, sondern können die gesamte Genexpression verschieben.
Für Schule, Studium und Labor würde ich mir vor allem drei Prüfsteine merken: Ist die Vorstufe vollständig gespleißt, sind die Enden korrekt verarbeitet und passt das Modifikationsmuster zur biologischen Aufgabe der RNA? Wer diese Fragen sauber beantwortet, versteht diesen Reifungsweg nicht nur als biochemischen Ablauf, sondern als zentrales Steuerungselement der Zelle. Genau deshalb bleibt das Thema weit mehr als ein Kapitel der Molekularbiologie - es ist ein Schlüssel zum Verständnis von Genregulation insgesamt.